This was originally written as an Answer to a Question posted to Scientific American Online; however, as what they published was considerably shorter and simpler than what I wrote, I shall post the [now updated] original here.
The answer to this question is not simple, because, while viruses all share the characteristics of being obligate intracellular parasites which use host cell machinery to make their components which then self-assemble to make particles which contain their genomes, they most definitely do not have a single origin, and indeed their origins may be spread out over a considerable period of geological and evolutionary time.
Viruses infect all types of cellular organisms, from Bacteria through Archaea to Eukarya; from E. coli to mushrooms; from amoebae to human beings – and virus particles may even be the single most abundant and varied organisms on the planet, given their abundance in all the waters of all the seas of planet Earth. Given this diversity and abundance, and the propensity of viruses to swap and share successful modules between very different lineages and to pick up bits of genome from their hosts, it is very difficult to speculate sensibly on their deep origins – but I shall outline some of the probable evolutionary scenarios.
The graphic depicts a possible scenario for the evolution of viruses: “wild” genetic elements could have escaped, or even been the agents for transfer of genetic information between, both RNA-containing and DNA-containing “protocells”, to provide the precursors of retroelements and of RNA and DNA viruses. Later escapes from Bacteria, Archaea and their progeny Eukarya would complete the virus zoo.
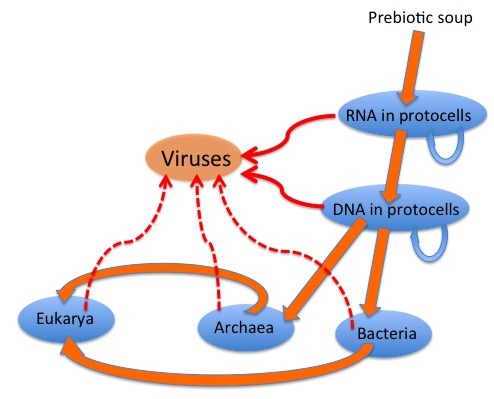
It is generally accepted that many viruses have their origins as “escapees” from cells; rogue bits of nucleic acid that have taken the autonomy already characteristic of certain cellular genome components to a new level. Simple RNA viruses are a good example of these: their genetic structure is far too simple for them to be degenerate cells; indeed, many resemble renegade messenger RNAs in their simplicity.
RdRp cassettes and virus evolution

RNA virus supergroups and RdRp and CP cassettes
What they have in common is a strategy which involves use of a virus-encoded RNA-dependent RNA polymerase (RdRp) or replicase to replicate RNA genomes – a process which does not occur in cells, although most eukaryotes so far investigated do have RdRp-like enzymes involved in regulation of gene expression and resistance to viruses. The surmise is that in some instances, an RdRp-encoding element could have became autonomous – or independent of DNA – by encoding its own replicase, and then acquired structural protein-encoding sequences by recombination, to become wholly autonomous and potentially infectious.
A useful example is the viruses sometimes referred to as the “Picornavirus-like” and “Sindbis virus-like” supergroups of ssRNA+ viruses, respectively. These two sets of viruses can be neatly divided into two groups according to their RdRp affinities, which determine how they replicate. However, they can also be divided according to their capsid protein affinities, which is where it is obvious that the phenomenon the late Rob Goldbach termed “cassette evolution” has occurred: some viruses that are relatively closely related in terms of RdRp and other non-structural protein sequences have completely different capsid proteins and particle morphologies, due to acquisition by the same RdRp module of different structural protein modules.
Given the very significant diversity in these sorts of viruses, it is quite possible that this has happened a number of times in the evolution of cellular organisms on this planet – and that some single-stranded RNA viruses like bacterial RNA viruses or bacteriophages and some plant viruses (like Tobacco mosaic virus, TMV) may be very ancient indeed.
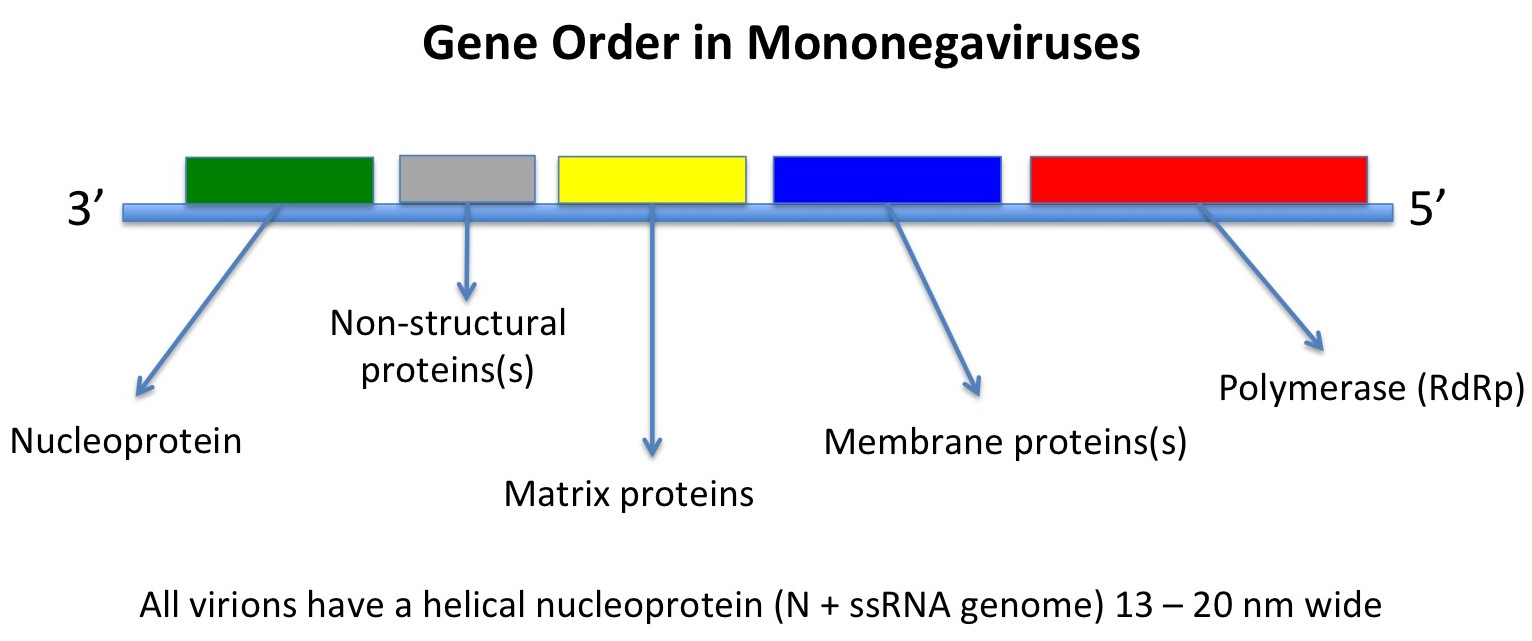
However, other ssRNA viruses – such as the negative sense mononegaviruses, Order Mononegavirales, which includes the families Bornaviridae, Rhabdoviridae, Filoviridae and Paramyxoviridae, represented by Borna disease virus, rabies virus, Zaire Ebola virus, and measles and mumps viruses respectively – may be evolutionarily much younger. In this latter case, the viruses all have the same basic genome with genes in the same order and helical nucleocapsids within differently-shaped enveloped particles.
Their host ranges also indicate that they originated in insects: the ones with more than one phylum of host either infect vertebrates and insects or plants and insects, while some infect insects only, or only vertebrates – indicating an evolutionary origin in insects, and a subsequent evolutionary divergence in them and in their feeding targets.

HIV: a retrovirus
The Retroid Cycle
The ssRNA retroviruses – like HIV – are another good example of possible cell-derived viruses, as many of these have a very similar genetic structure to elements which appear to be integral parts of cell genomes – termed retrotransposons – and share the peculiar property of replicating their genomes via a pathway which goes from single-stranded RNA through double-stranded DNA (reverse transcription) and back again, and yet have become infectious. They can go full circle, incidentally, by permanently becoming part of the cell genome by insertion into germ-line cells – so that they are then inherited as “endogenous retroviruses“, which can be used as evolutionary markers for species divergence.

The Retroid Cycle
Indeed, there is a whole extended family of reverse-transcribing mobile genetic elements in organisms ranging from bacteria all the way through to plants, insects and vertebrates, indicating a very ancient evolutionary origin indeed – and which includes two completely different groups of double-standed DNA viruses, the vertebrate-infecting hepadnaviruses or hepatitis B virus-like group, and the plant-infecting badna- and caulimoviruses.
Metaviruses and pseudoviruses
These are two families of long terminal repeat-containing (LTR) retrotransposons, with different genetic organisations.
Members of family Pseudoviridae, also known as Ty1/copia elements, have polygenic genomes of 5-9 kb ssRNA which encode a retrovirus-like Gag-type protein, and a polyprotein with protease (PR), integrase (IN) and reverse transcriptase / RNAse H (RT) domains, in that order. While some members also encode an env-like ORF, the 30-40 nm particles that are an essential replication intermediate have no envelope or Env protein. They are not infectious. Host species include yeasts, insects, plants and algae.
Metaviruses – family Metaviridae – are also known as Ty3-gypsy elements, and have ssRNA genomes of 4-10 kb in length. They replicate via particles 45-100 nm in diameter composed of Gag-type protein, and some species have envelopes and associated Env proteins. Gene order in the genomes is Gag-PR-RT-IN-(Env), as for retroviruses. One virus – Drosophila melanogaster Gypsy virus – is infectious; however, as for pseudoviruses, most are not. The genomes have been found in all lineages of eukaryotes so far studied in sufficient detail.
Both pseudovirus and metavirus genomes are clearly related to classic retroviruses; moreover, RT sequences point to metavirus RTs being most closely related to plant DNA pararetrovirus lineage of caulimoviruses. This gives rise to the speculation that pseudoviruses and metaviruses have a common and ancient ancestor – and that two different metavirus lineages gave rise to retroviruses and caulimoviruses respectively.
All of these cellular elements and viruses have in common a “reverse transcriptase” or RNA-dependent DNA polymerase, which may in fact be an evolutionary link back to the postulated “RNA world” at the dawn of evolutionary history, when the only extant genomes were composed of RNA, and probably double-stranded RNA. Thus, a part of what could be a very primitive machinery indeed has survived into very different nucleic acid lineages, some viral and many wholly cellular in nature, from bacteria through to higher eukaryotes.
The possibility that certain non-retro RNA viruses can actually insert bits of themselves by obscure mechanisms into host cell genomes – and afford them protection against future infection – complicates the issue rather, by reversing the canonical flow of genetic material. This may have been happening over aeons of evolutionary time, and to have involved hosts and viruses as diverse as plants (integrated poty– and geminivirus sequences), honeybees (integrated Israeli bee paralysis virus) – and the recent discovery of “…integrated filovirus-like elements in the genomes of bats, rodents, shrews, tenrecs and marsupials…” which, in the case of mammals, transcribed fragments “…homologous to a fragment of the filovirus genome whose expression is known to interfere with the assembly of Ebolavirus”.
Rolling circle replication
There are also obvious similarities in mode of replication between a family of elements which include bacterial plasmids, bacterial single-strand DNA viruses, and viruses of eukaryotes which include geminiviruses and nanoviruses of plants, parvoviruses of insects and vertebrates, and circoviruses and anelloviruses of vertebrates.

Geminivirus particle
These agents all share a “rolling circle” DNA replication mechanism, with replication-associated proteins and DNA sequence motifs that appear similar enough to be evolutionarily related – and again demonstrate a continuum from the cell-associated and cell-dependent plasmids through to the completely autonomous agents such as relatively simple but ancient bacterial and eukaryote viruses.

Big DNA viruses

Mimivirus particle, showing basic structure
However, there are a significant number of viruses with large DNA genomes for which an origin as cell-derived subcomponents is not as obvious. In fact, one of the largest viruses yet discovered – mimivirus, with a genome size of greater than 1 million base pairs of DNA – have genomes which are larger and more complex than those of obligately parasitic bacteria such as Mycoplasma genitalium (around 0.5 million), despite their sharing the life habits of tiny viruses like canine parvovirus (0.005 million, or 5000 bases).
Mimivirus has been joined, since its discovery in 2003, by Megavirus (2011; 1.2 Mbp) and now Pandoravirus (2013; 1.9 -2.5 Mbp).
The nucleocytoplasmic large DNA viruses or NCLDVs – including pox-, irido-, asfar-, phyco-, mimi-, mega- and pandoraviruses, among others – have been grouped as the proposed Order Megavirales, and it is proposed that they evolved, and started to diverge, before the evolutionary separation of eukaryotes into their present groupings.
It is a striking fact that the largest viral DNA genomes so far characterised seem to infect primitive eukaryotes such as amoebae and simple marine algae – and they and other large DNA viruses like pox- and herpesviruses seem to be related to cellular DNA sequences only at a level close to the base of the “tree of life”.

Variola virus, the agent of smallpox. Image courtesy Russell Kightley Media.
This indicates a very ancient origin or set of origins for these viruses, which may conceivably have been as obligately parasitic cellular lifeforms which then made the final adaptation to the “virus lifestyle”.
However, their actual origin could be in an even more complex interaction with early cellular lifeforms, given that viruses may well be responsible for very significant episodes of evolutionary change in cellular life, all the way from the origin of eukaryotes through to the much more recent evolution of placental mammals. In fact, there is informed speculation as to the possibility of viruses having significantly influenced the evolution of eukaryotes as a cognate group of organisms, including the possibility that a large DNA virus may have been the first cellular nucleus.
In summary, viruses are as much a concept as a unitary entity: all viruses have in common, given their polyphyletic origins, is a base-level strategy for replicating their genomes. Otherwise, their origins are possibly as varied as their genomes, and may remain forever obscure.
I am indebted to Russell Kightley for use of his excellent virus images.
Updated 12th August 2015
Tags: circovirus, DNA, Evolution, geminivirus, herpesvirus, mammals, mimivirus, poxvirus, retrotransposon, retrovirus, RNA, rolling circle, virus

2 October, 2008 at 14:18 |
[…] samples, or even blood of pandemic survivors to speculate on the origins of specific viruses, of viruses generally, or on the nature of old pandemic strains. Now HIV falls under the spotlight – again – as the […]
LikeLike
11 September, 2015 at 09:03 |
are there any study which postulates the exact era of appearance and evolution of viruses? are there any paleantological evidence of HIV?
LikeLike
20 November, 2008 at 16:30 |
[…] large DNA viruses – pox-, irido-, herpes-, asfar-, mimi- and baculoviruses and their ilk – have a deep and possibly complex evolutionary history, and there is considerable evidence to suggest long histories of co-speciation […]
LikeLike
21 July, 2009 at 15:55 |
Why do virus originate before eukaryotes and prokaryotes? Are there any reasons or theories for this?
LikeLike
21 July, 2009 at 16:20 |
Hi Joleen: well, there is no evidence that they did…but almost certainly viruses were there when the first cells were getting going, meaning before eukaryotes – and they may even have helped give rise to eukaryotes, if you believe the “captured virus” scenario for the origin of the nucleus.
LikeLike
13 June, 2010 at 22:02 |
Well, there is some indirect evidence. There are structural similarities between the capsids of adenovirus (that infect mammals and birds), and some bacteriophage that infect eubacteria (plain ordinary bacteria if you like) and others that infect archaea (a totally different class of bacteria). One way to explain this observation is to propose that the ancestor of these three different viruses was around before the eubacteria/archaebacterai/eukaryote split.
Some similar parallels can be found between herpesviruses, that infect a wide range of vertebrate and invertebrate hosts, and the tailed phages (like T4, or lambda) that infect bacteria.
With these examples, it’s difficult to be sure whether the similarities are due to common ancestry, or to gene transfer between viruses infecting different hosts. However, since these similarites are only visible in 3-d structures, not in sequences, they are probably extremely ancient.
DMc
LikeLike
1 August, 2009 at 18:36 |
Hi Ed: Thank you for the reply. I have a better understanding of the virus now. Thank you.
LikeLike
2 April, 2010 at 16:21 |
Just when you thought that Mimivirus had enough genes, we find 75 more!
http://454.com/about-454/news/index.asp?display=detail&id=144
LikeLike
3 April, 2010 at 11:43 |
Incroyable…! Thanks; I am sure there is an enormous amount to learn from these monster viruses.
LikeLike
21 April, 2011 at 20:08 |
[…] ViroBlogy праз kwout […]
LikeLike
17 June, 2011 at 07:56 |
Hi Ed,
Great blog, thanks for all of the fascinating posts.
In this article you mention: “The possibility that certain non-retro RNA viruses can actually insert bits of themselves by obscure mechanisms into host cell genomes – and afford them protection against future infection – complicates the issue rather, by reversing the probable flow of genetic material!”
However, the link is broken! I can easily find my way back to http://www.microbiologybytes.com but am having trouble locating your source for this information. Could you please help me find locate it, or point me in the right direction to find it elsewhere.
Thanks,
Chad
LikeLike
17 June, 2011 at 08:08 |
Well, that did not take as long as expected!
Assuming this is the page you linked to in the original article above, this will fix the broken link: http://www.microbiologybytes.com/blog/2007/09/14/dont-look-now-theyre-in-your-genes/
Thanks for the great blog!
LikeLike
17 June, 2011 at 10:30 |
Chad: thank you! Link fixed and I tidied up the font mish-mash too.
LikeLike
28 September, 2011 at 12:32 |
[…] Virus origins: from what did viruses evolve or how did they initially arise? […]
LikeLike
28 September, 2011 at 13:54 |
[…] have updated the blog on virus origins quite considerably – new pictures, more detail, more speculation! Pathways on information […]
LikeLike
29 September, 2011 at 09:45 |
[…] Virus origins: from what did viruses evolve or how did they initially arise? […]
LikeLike
11 October, 2011 at 10:54 |
[…] those earlier papers appears to be borne out here. Namely, and quite important when considering both viral and cellular origins, is further evidence that very large viral genomes do not seem to have evolved by extensive […]
LikeLike
12 December, 2011 at 18:44 |
On The RNAs, Life’s Primal ORGANISMS…:
The DNA and RNA genomes are ORGANISMS evolved by life’s primal ORGANISMS, the RNAs, and so are all cells…
(Extend evolution way down to genes, life’s base ORGANISMS. Culture modifies genetics, not vice versa…)
Pavlov’s Smile: RNAs Are Earth’s Primal Organisms
Culture>genes>addiction (2 July 2009)
http://universe-life.com/2011/09/24/pavlovs-smile/
Why Pavlov smiled in 2008?
Pavlov demonstrated effecting placebo phenomena in multi celled organisms by manipulation of their drives-reactions. Now placebo and imagination phenomena are demonstrated also in Earth’s smallest, base organisms, in the genes and genomes of multi-celled organisms, in our primal 1st stratum and 2nd stratum base organisms.
A very good reason to smile.
Now an interesting chain is exposed to our view, the Genes-Virtual Reality Chain, a most intriguing cultural evolution chain extending from the genesis of our genes to nowadays, throughout life, a virtual reality existence, and by virtual reality phenomena, exploitations and manipulations.
Dov Henis (comments from 22nd century)
http://universe-life.com
PSW1:
From “Life Genesis From Aromaticity/H-Bonding”
http://universe-life.com/2011/09/30/earthlife-genesis-from-aromaticityh-bonding/
Natural selection is E (energy) temporarily constrained in an m (mass) format.
Natural selection is a universal ubiquitous trait of ALL mass spin formats, inanimate and animate.
Life began/evolved on Earth with the natural selection of inanimate RNA, then of some RNA nucleotides, then arriving at the ultimate mode of natural selection, self-replication.
PS2:
If I told you once, I told you a million times: It’s The Horses Pulling, Not The Wagon Pushing
http://universe-life.com/2011/08/26/on-culture-and-genetics-horses-and-wagon/
LikeLike
29 January, 2012 at 12:46 |
Radiation can damage many cellular targets, and the result is that ” molecular parasites ” or viruses.
LikeLike
29 January, 2012 at 20:18 |
This may have been, once or twice upon a time, the origin of one or more viruses – but is hardly a regular thing!
LikeLike
9 February, 2012 at 20:20 |
I have a kind of unrelated question, so we are not sure from what viruses evolved, but do we know to what organisms did viruses evolve? did they evolve into bacterias?
LikeLike
16 April, 2012 at 04:00 |
New Era For Science Including Genomics ???
From: Dov Henis
Sent: Friday, April 13, 2012 10:43 PM
To: genome biologists
Subject: A new era for science including genomics ??? Please examine carefully…
Yesterday, SN , after many years of refusing my similar postings, SN posted my following statement-comment:
http://www.sciencenews.org/index/generic/activity/view/id/339659/title/Fruit_fly_biorhythms_differ_indoors_and_out
Biorhythms Schmiorythms
Circadian Schmircadian sleep origin?
Life sleeps because RNAs genesized, evolved from inanimate nucleotides into self-replicating nucleotides, organisms, of course long before metabolism evolved. They were then active ONLY during sunlight hours. Thus sleep is inherent for RNAs, even though, being ORGANISMS, they later adapted to when/extent sleep times are feasible just as we adapt to jetlag or night work time.
Dov Henis (comments from 22nd century)
Apr. 12, 2012 at 9:10am
===========================
From: Dov Henis
Sent: Saturday, April 14, 2012 9:05 AM
To: genome biologists
Subject: FW: A new era for science including genomics ??? Please examine carefully…
Unbelievable?! Here’s another one…
From: Dov Henis
Sent: Sunday, April 08, 2012 9:06 PM
To: ‘editors@sciencenews.org’
Subject: On Pavlov and genes…
Fatty Diet Leads To Fat-Loving Brain Cells
http://www.sciencenews.org/view/generic/id/339458/title/Fatty_diet_leads_to_fat-loving_brain_cells
Learn from Pavlov:
Fatty diet lead to fat-loving RNA-nucleotides genes, Earthlife base primal ORGANISMS.
Dov Henis (comments from 22nd century)
http://universe-life.com/2011/12/22/rnas-are-earths-primal-organisms/
===================
Since the above two statements are basis for the following statement, may it also soon pass the SN “peer review”… ?!
USA Science? Re-Comprehend Origins And Essence
• Higgs Particle? Dark Energy/Matter? Epigenetics? All YOK!
• Earth-life is just another, self-replicating, mass format.
• All mass formats follow natural selection, i.e. intake of energy or their energy taken in by other mass formats.
• Evolution Is The Quantum Mechanics Of Natural Selection.
• Quantum mechanics are mechanisms, possible or probable or actual mechanisms of natural selection.
• Life’s Evolution is the quantum mechanics of biology.
• Every evolution, of all disciplines, is the quantum mechanics of the discipline’s natural selection.
See:
Update Concepts-Comprehension…
http://universe-life.com/2011/12/13/21st-century-science-whence-and-whither/
Earth life genesis from aromaticity-H bonding
http://universe-life.com/2011/09/30/earthlife-genesis-from-aromaticityh-bonding/
Universe-Energy-Mass-Life Compilation
http://universe-life.com/2012/02/03/universe-energy-mass-life-compilation/
Seed of human-chimp genome diversity
http://universe-life.com/2011/07/10/seed-of-human-chimp-genomes-diversity/
============
Respectfully,
Dov Henis (comments from 22nd century)
LikeLike
9 May, 2012 at 11:24 |
Reblogged this on Outline Science.
LikeLike
11 September, 2013 at 16:30 |
[…] I can finally update a speculative "modular evolution" diagram I constructed a while back, with actual evidence: https://rybicki.wordpress.com/2008/03/19/from-what-did-viruses-evolve-or-how-did-they-initially-arise… […]
LikeLike
11 September, 2015 at 08:56 |
very concise & informative article
LikeLike
11 September, 2015 at 08:59 |
Reblogged this on whistlingintherain and commented:
very concise and apt article for students interested in virology
LikeLike
14 February, 2016 at 06:35 |
Sadahatama Oba Mage
Virus origins: from what did viruses evolve or how did they initially arise? | ViroBlogy
LikeLike
31 August, 2016 at 16:16 |
[…] have been transmitted by arthropods. Because that’s the link between many of these viruses: an evolutionary history that involves plants and arthropods, or arthropods and other animals, at an early stage of life on land. Because that’s all there […]
LikeLike