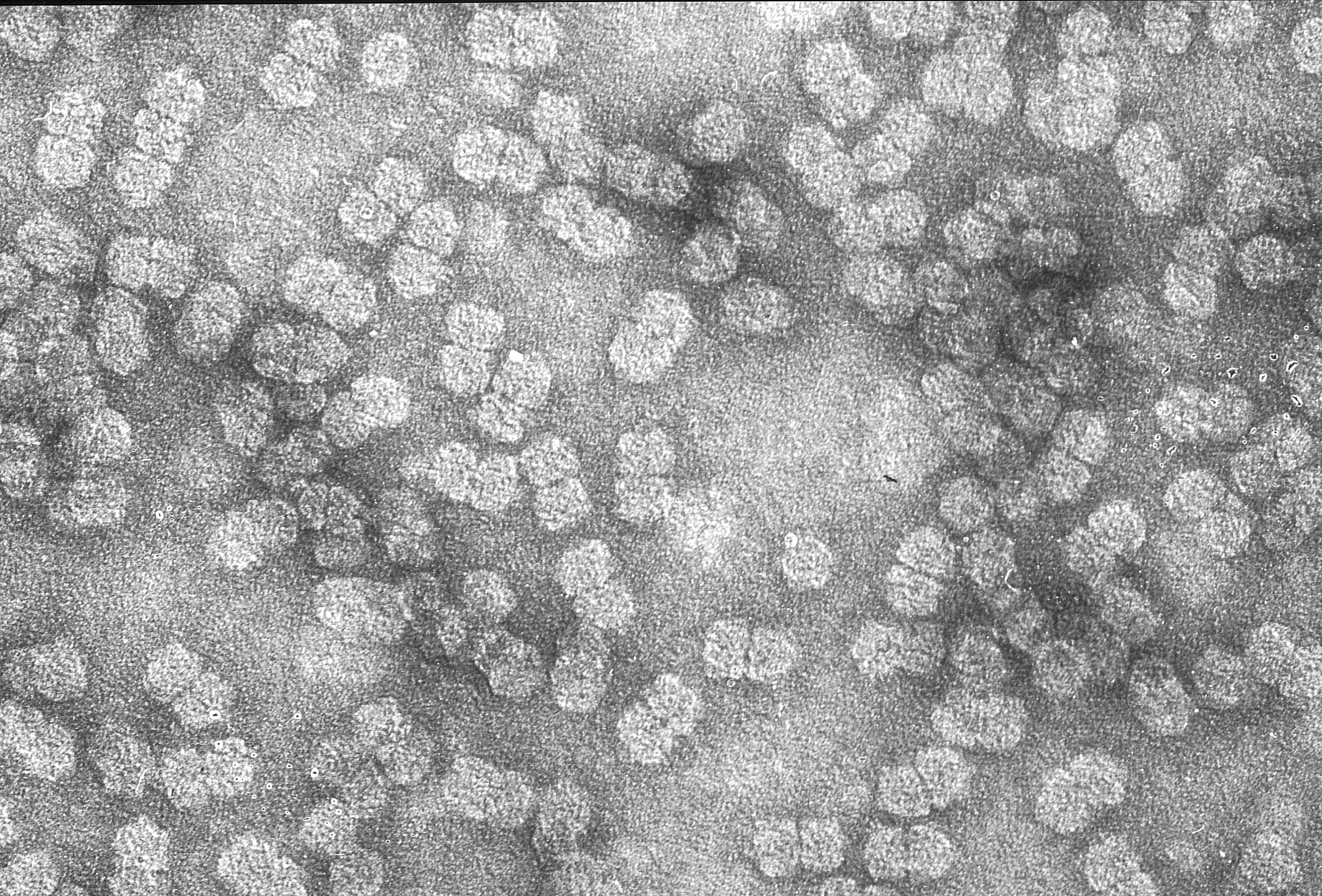
A Maize streak virus particle: characteristic doubled icosahedron containing a single ssDNA
Graphic by Russell Kightley Media
I blogged on this paper from our group over at Virology News, but then I decided to do it again here. Because, as I said there,
“This is a big deal: seriously. It’s the culmination of some 24 years of involvement by my lab in engineering resistance in maize, and is the latest effort on top of of one unsuccessful and one partially successful construction by three top-class researchers in that time.”
…and I don’t think I did justice at the time to acknowledging the amount of effort it has taken to get to this point – which has occupied nearly 30 years of my life, and a considerable fraction of the working life of many others. So here’s something of a chronicle of how we got here from there with some explanation of the very cool science behind it, and acknowledgement of a very valuable friendship between us and colleagues in Australia, without whom this would not have happened.
Maize streak virus is, of course, possibly the first virus described from Africa, and one famous enough to have its own web page, complete with the Proceedings of the First (and last) International Maize Streak Disease Symposium held in Hazyview in South Africa in 1997. It is a ssDNA virus with a 2.7 kb circular genome that is encapsidated in unique geminate particles, is obligately transmitted by the leafhopper Cicadulina mbila Naude, and is probably the worst viral pathogen of maize in Africa.
The Maize streak virus investigations that led to this latest development were started by my retired colleague and former PhD supervisor Prof Barbara von Wechmar, back in the 1960s at the University of Stellenbosch. Barbara it was who isolated MSV out of naturally infected maize, and developed home-grown methods of propagating it in sweetcorn via viruliferous Cicadulina mbila leafhoppers – and more importantly, getting clean leafhopper colonies so that new isolates could be studied.
She brought this knowledge with her to UCT when she came here with Marc van Regenmortel as the new Professor of Microbiology, and the leafhopper work and virus isolates puttered on in the background, with a couple of interesting papers coming out. The first was a collaborative work with the legendary Bob (RG) Milne, who came here on a sabbatical from the Istituto di Fitovirologia Applicata del CNR in Torino, Italy, and revolutionised our electron microscopy techniques by use of uranyl acetate instead of phosphotungstate, which meant for the first time we could see MSV particles. This got presented at an International Maize Virus Disease Colloquium and Workshop in Wooster, Ohio in 1982, the Proceedings of which seem to be available here.
VON WECHMAR, M.B., & MILNE, R.G. (1983). Purification and serology of a South African isolate of maize streak virus. pp. 164-166 In: Proc. Internat. Maize Virus Dis. Colloquium and Workshop. University of Ohio Press, Wooster

Maize streak virus particles isolated from maize, photographed by Robert G Milne in Cape Town
Another paper in J Gen Virol from 1983 was a fascinating account of using “electroinfection” to infect maize plants with MSV, published with Dr Alfred Polson: he was a quirky and idiosyncratic physical biochemist-cum-virologist who had done a PhD with The Svedberg and Ole Lamm in Uppsala in the 1930s, headed a Virus Research Unit at UCT Medical School, then retired to our Department to basically play with physical and serological techniques. He liked nothing better than to hook plants up to kilovolt power packs, and to do large-scale electrophoresis of proteins and viruses in convoluted home-made glass contraptions – and to pull us in to help.
Also in the early 1980s, when molecular microbiology belatedly dawned in South Africa, a couple of DNA-wise colleagues and I thought it would be a great idea to sequence the MSV genome, given that it had been shown in 1977 in a Nature paper by Bryan Harrison and colleagues to be circular ssDNA. Sadly, this never really got past the talking and wasting virus sample stage by Frank Robb and Ralph Kirby before two papers were published on the sequence of MSVs in 1984 – with the second, by Phil Mullineaux and colleagues from the John Innes Institute, drily pointing out that the first, by Steve Howell, had the sequence of the complementary, rather than the genomic strand of DNA.
I decided in 1985, after getting a PhD on a biophysical and serological investigations of small grain viruses in 1984, to change my skill set during a three-month academic leave in Belgium by learning molecular cloning and DNA handling techniques. Thanks to the recommendation of Marc van Montagu, and the teachings of the good folks at Plant Genetic Systems in Gent (Jan Leemans and Herman Hofte, I still owe you B-), this was achieved – and the first thing I did on my return was to recruit an able Honours student in the person of Bev Clarke, cosupervised by Ralph Kirby for the DNA expertise, to clone and sequence local MSV strains that were still being maintained by Barbara von Wechmar. Bev worked through into a Masters degree on cloning and restriction mapping of three maize isolates of MSV – sequencing was still a bit complicated and expensive in those days – and we managed to publish two babck-to-back papers out of her work, in 1989. The first was an account of the propagation, isolation, cloning and mapping, and we in fact got the cover of that issue.
 The second was the start of what became a really interesting sideline for me, in evolutionary studies on viruses – because Ralph managed to use the maps we generated from our viruses and from sequenced isolates to estimate sequence divergence. It really is quite amusing, in this era of rapid and metagenomic sequencing, to read what we thought we had accomplished at the time, given that my team has just sequenced some ten geminiviruses as part of a BSc third year project:
The second was the start of what became a really interesting sideline for me, in evolutionary studies on viruses – because Ralph managed to use the maps we generated from our viruses and from sequenced isolates to estimate sequence divergence. It really is quite amusing, in this era of rapid and metagenomic sequencing, to read what we thought we had accomplished at the time, given that my team has just sequenced some ten geminiviruses as part of a BSc third year project:
“The aligned restriction endonuclease maps of three sequenced maize streak virus isolates, three restriction-mapped southern African maize streak virus isolates, and two other sequenced geminiviruses were used as a means of calculating the sequence divergence between these viruses. The degree of divergence was used to construct a phylogenetic tree for the viruses; this tree agrees well with predictions from sequence comparisons, and so the method can be used to study the relationship of geminivirus isolates without the labor and expense of sequencing each one. [my emphasis]”
On the strength of this and other work, I was prompted in 1988 to write a bold (and naive) lamppost-marking type of article on “Maize streak virus: an African pathogen come home?”, which I commemorated 25 years later here in ViroBlogy. I cringed a little then and again now, seeing what we thought was so cool at the time. Ah, youth…B-)
Bev was succeeded as “the” student in my lab by Fiona Tanzer (then Hughes), who took over the MSV and other geminivirus work with verve and flair. She produced a number of papers during her PhD, starting with an offering on “A rapid technique for typing streak virus isolates using a panel of differential hosts” at a South African Maize Breeding Symposium in 1990, and going on to characterise Sugarcane streak mastrevirus (SSV) from Natal as a distinct virus in 1991, by RE mapping, Southern hybridisation and partial sequence analysis; “Genome Typing of Southern African Subgroup-1 Geminiviruses” by the same techniques in 1992, and graduating to “Complete nucleotide sequence of sugarcane streak Monogeminivirus” in 1993.
Meantime, in what turned out to be my last stint of concentrated laboratory work, I had been investigating the potential of PCR for both detection and differentiation of mastreviruses – which led to a little detour into papillomavirology to apply the technique, which ended up being a complete change in direction, and a whole new career in vaccinology. I used the primer design experience and PCR results I got then as the basis of an online teaching module on PCR, which is still available – and for which I actually get citations.

Degenerate Primer PCR for Mastrevirus Detection
But I digress: Fiona went on after her PhD to work as a postdoc with me and Jennifer Thomson, with whom I had been collaborating since she arrived at UCT in 1988 in the area of plant genetic engineering, on an insanely ambitious project to engineer transgenic resistance to MSV in maize. This had in fact started in 1990, when I visited Bill Gordon-Kamm at DeKalb Genetics in Connecticut while on sabbatical at Cornell in 1990-1991: I got a suspension culture of Black Mexican Sweet (BSM) maize cells from them for us to practice biolistic techniques on with our newly-acquired BioRad helium gene gun, as well as a collaboration agreement which helped us immensely in our maize transformation and regeneration work. Sandy Lennox it was who got that side of things working, to the extent of regenerating viable plants from supposedly non-regenerable BMS cells – which laid the foundation for all of our subsequent work in this area, using HiII embryogenic callus cultures.
Fiona, meanwhile, had managed to both clone and sequence a moderately severe isolate of MSV from Komatipoort (MSV-Kom), and then also make it agroinfectious as a partially dimeric clone in Agrobacterium tumefaciens, following the landmark 1987 example of Nigel Grimsley and colleagues in Basel. This provided the other half of the toolkit, as we now had a means of reliably testing regenerated maize with a MSV isolate of known virulence, available for infection as a clone rather than transmitted via leafhoppers.
Fiona went on to make and test antisense RNA-expressing constructs from the Rep gene of MSV-Kom, as I had been heavily influenced by a 1991 Keystone Symposium on Antisense RNA Technology I went to while on sabbatical – and reported on here – and was convinced this was the way to go.
Inevitably, as determined by exhaustive experimentation from laboriously regenerated plants, this turned out not to be true: there was no obvious protection of the highly susceptible HiII by any of the constructs. Fiona went off to have a baby – her second while working with me – and the MSV effort stalled for a while. Barbara retired, and it fell to me to maintain leafhopper colonies and MSV and other mastrevirus isolates – which, very fortunately, we had begun routinely cloning and making agroinfectious, via a very able cohort of students led by Wendelin Heribert “Popeye” Schnippenkoetter, who perfected the art of the “1.1-mer” infectious clone as well as finally publishing the MSV-Kom sequence.
The value of international networking in geminivirology became apparent in 1994, while all this this was happening, when I went to the first International Geminivirus Symposium in Almeria, Spain. There I re-made the acquaintance of Doug Maxwell, who I had met in Ithaca in 1991 when he spoke on using PCR to detect begomoviruses. I also recall performing “Born to be Wild” in a hotel karaoke bar with a scratch band of geminivirologists, but we shall speak no further of this.

Doug and I corresponded until the next Symposium he organised in Puerto Rico in 1998, when we heard his PhD student Steve Hanson present on the use of trans-dominant Rep gene mutants to inhibit the replication of Bean golden mosaic begomovirus in transient assays in cultured bean cells. They sent us the PhD thesis to use as a reference document; the work was eventually published in 1999.
This kicked our efforts into a higher gear, given that we now had an exemplar in another albeit VERY distantly related geminivirus. We also had a very bright new PhD student from Zimbabwe, Tichaona Mangwende, as the perfect guinea pig for investigation of dominant negative Rep mutants, and off we set again. Tich quickly made a number of site-directed mutants by very ingenious methods, and we then came up to the hurdle of how to test them. We had had in mind that this would be done in regenerated plants; however, it was quickly apparent that this would stretch his project out for several years longer than was feasible.
Here it was that the value of having students working in related projects was shown: after 1996 I had received expanded funding due to a favourable Foundation for Research Development rating, and actually had a lab full of students all working on geminiviruses. Fortuitously, Kenneth Palmer had been doing a project on exploring the potential of MSV-based constructs as recombinant expression vectors for use in maize, and had worked out a suite of techniques for bombarding BMS suspension-cultured cells and assaying for virus genome replication and protein expression. Even more fortuitously, Janet Willment had been investigating the minimal cis-acting control regions for replication of MSV and their sequence specificity, and following Kenneth’s example, had set up exactly the right system for biolistic introduction of DNA constructs into suspension cultured cells as well as quantitative PCR for assay of replication, that Tich required for his work. He was able to test three different constructs in conjunction with partial dimers of MSV-Kom by transient expression in biolistically transformed cells, and prove that they significantly inhibited MSV genome replication. He got a great PhD, and has gone on to good things in the agricultural biotech sector in SA.
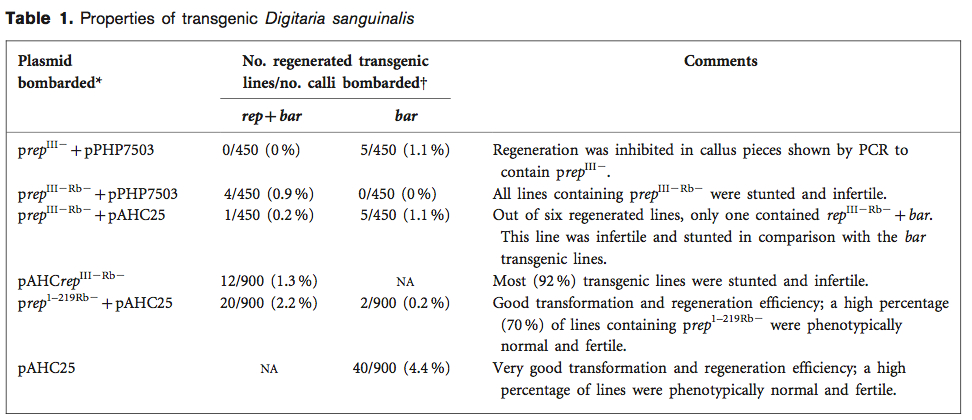
The stage was now set and dressed for the final act: this was the introduction of Dionne Miles, now Shepherd, into the continuation of this project into whole plant testing. Here also was another example of fortuitous cross-bleed between projects: it happened that Kenneth Palmer had been helping Jennifer Thomson with a PhD student who was working on a difficult project to do with regeneration of cereals from anther culture. He helped Wusi develop a system for transformation and regeneration of a model grass species, Digitaria sanguinalis, originally sourced from the flowerbed next to the UCT Sports Centre. As it happened, some regenerated Digitaria was in a plant room that had escaped viruliferous leafhoppers in it – and got infected with MSV-Kom, and showed splendid streak symptoms as well as stunting. As it was easier to transform than maize, could be grown as a perennial by simply cutting it back, and went to seed inside six weeks, it was obvious that we had a wonderful model plant for MSV resistance testing. Another vital cog in what was becoming a complicated machine was my student Darrin Martin, who had developed a truly wonderful image processing-based quantitative symptom assessment tool as a central part of his PhD project on the determinants of pathogenicity in MSV. This also proved vital in future work on accurate determination of the degree of resistance of regenerated maize.

Dionne started what was to become her whole professional life to date in the late 1990s, by modifying Tich’s clones and testing them in maize cells for efficacy, and then introducing them into D sanguinalis cells for regeneration. The value of the strategy was quickly apparent, when she showed that the best constructs for transient inhibition of MSV replication also either prevented regeneration of plants completely, or produced a very aberrant and infertile phenotype.

She was left with one – a truncated Rb- mutant Rep, rep1-219Rb- – that allowed regeneration of normal fertile plants and inhibited virus replication in transient assays, that has formed the basis of most of the work since. This includes the publication of her development of MSV-resistant transgenic fertile maize as “Maize streak virus-resistant transgenic maize: a first for Africa” in the Plant Biotechnology Journal in 2007, and getting us another cover.


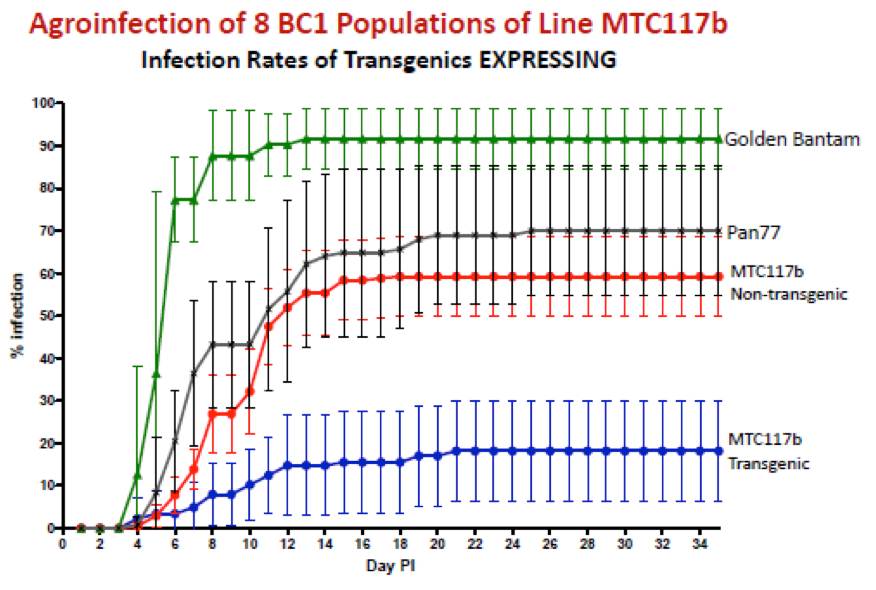
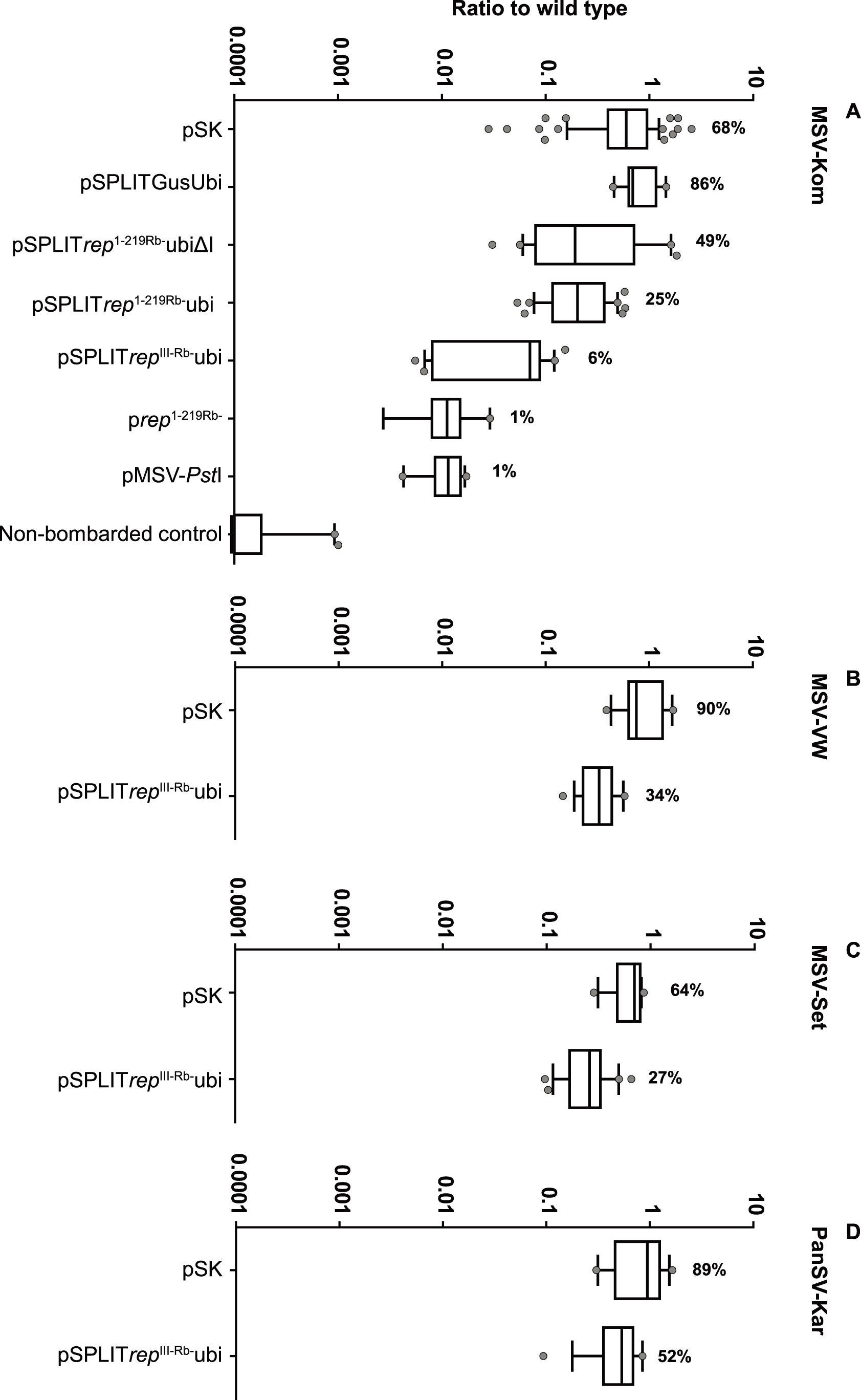
What followed this was a long, painstaking grind by Dionne and team, and notably Marian Bezuidenhout who did most of the transformation and regeneration, in making as many transgenic lines as possible to provide to our maize seed producing industry partner Pannar Pty Ltd for introgression of the transgene into their elite breeding lines. There followed much assessment of symptom development in greenhouse-tested plants, often grown from or descended from plantlets flown from our plant rooms to Greytown in an executive jet – along with boxes of wine, it does need to be said. This has produced analyses that look like this: the product of more patient work than I think I would be capable of, and a really good example of how one should do this sort of work. With repeats. Many, many repeats.

Dionne followed this up in 2011 with a second-generation product, with a paper on “A rep-based hairpin inhibits replication of diverse maize streak virus isolates in a transient assay“:
“After co-bombardment of cultured maize cells with each construct and an infectious partial dimer of the cognate virus genome (MSV-Kom), followed by viral replicative-form-specific PCR, it was clear that… the hairpin rep construct (pHPrepΔI(662)) completely inhibited MSV replication…[and] in addition, pHPrepΔI(662) inhibited or reduced replication of six MSV-A genotypes representing the entire breadth of known MSV-A diversity.”
This is also a big deal, as it represents another, alternative strategy to confer MSV resistance on maize, that confers wider resistance, and could potentially be stacked with the previous construct.
Then at last, we come to the present work – with a sense of the history behind it. The rationale for this was the following:
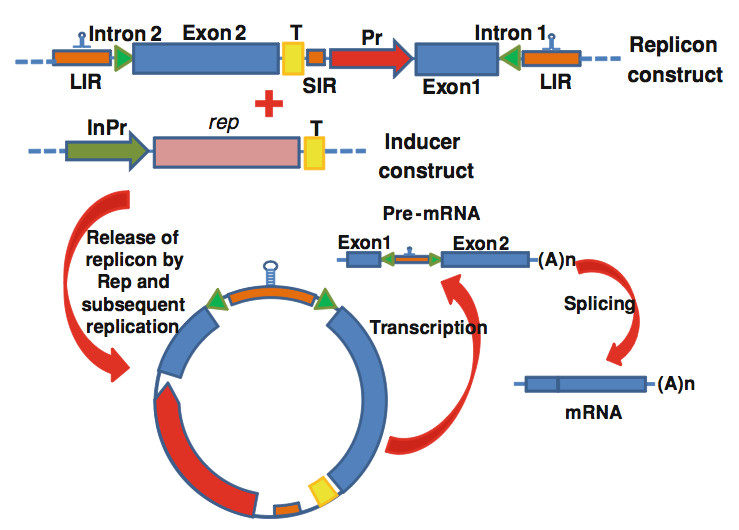
“While we have previously developed MSV-resistant transgenic maize lines constitutively expressing “dominant negative mutant” versions of the MSV Rep, the only transgenes we could use were those that caused no developmental defects during the regeneration of plants in tissue culture. A better transgene expression system would be an inducible one, where resistance-conferring transgenes are expressed only in MSV-infected cells. However, most known inducible transgene expression systems are hampered by background or “leaky” expression in the absence of the inducer. Here we describe an adaptation of the recently developed INPACT system to express MSV-derived resistance genes in cell culture.”
Backtracking slightly, this has been the product of another networking experience at a foreign conference: this time, between me and my by-then-old friend James Dale from Queensland University of Technology, at the “Virologica 2001” conference of the Brazilian Society for Virology in Caldas Novas, Brazil. Relaxing as invited speakers do, with a beer on a balcony, James said to me “Ed, mate, you’re going to kick yourself that you didn’t think of this!”, and proceeded to tell me about a geminivirus-based inducible expression system he and his group had just invented. I did. I also swore, loudly. I then got him to offer highly reasonable terms for us to use the technology, and promptly took the idea home. I have also described it in a recent review on ssDNA virus-derived plant expression vectors, so I take great glee in presenting my graphic here:

This has only been published recently, due to patent issues and its use in proprietary production – however, we got the full cooperation of James and Ben Dugdale of QUT early on, and
“…used a quantitative real-time PCR assay to show that one of these SGCs (pSPLITrepIII-Rb-Ubi) inducibly inhibits MSV replication as efficiently as does a constitutively expressed transgene that has previously proven effective in protecting transgenic maize from MSV. In addition, in our cell-culture based assay pSPLITrepIII-Rb-Ubi inhibited replication of diverse MSV strains, and even, albeit to a lesser extent, of a different mastrevirus species. The application of this new technology to MSV resistance in maize could allow a better, more acceptable product.”

Pretty good, you’d think? Also an excellent example of South-South collaboration, and investigations being set up as they should: by two people having a beer, at a conference. Thanks, James! Thanks Benno! The paper ends optimistically, with:
“Ultimately, the practicality of the SGCs described in this study will only be fully realised with the regeneration of phenotypically normal transgenic maize plants engineered to contain the SGC that are resistant/immune to MSV infection. To this end we have regenerated a number of transgenic maize lines containing a SGC capable of expressing the most effective Rep mutant, namely RepIII-Rb-. In contrast to lines constitutively expressing this mutant gene, SGC lines have produced T2generation offspring with normal phenotypes.
Considering that only one strain – MSV-A – causes severe disease in maize throughout the whole geographical range of MSV, and that all isolates so far discovered within this strain have a maximum divergence of only 4.62% at the nucleotide level, it is likely that this novel MSV-inducible resistance construct will be effective against the complete spectrum of severe maize streak disease-causing African MSVs.”
Sadly, and these promises notwithstanding, this has all come to an end, with the non-renewal of the funding – possibly as a result of the takeover of Pannar by Pioneer HiBred Intl, but for whatever reason, it marks literally the end of an era.
What the hell – we had fun. And maybe we can still sell it to someone.