I promised some time ago now to blog on the exciting topic of whether or not a plant virus is infecting honeybees – and here it is! I was also contacted by the legendary Dr Adrian Gibbs about this paper, because he has read this blog, so I am including a commentary from him as well.
A little while ago, Ji Lian Li and co-workers published a paper entitled “Systemic Spread and Propagation of a Plant-Pathogenic Virus in European Honeybees, Apis mellifera” in ASM’s Open Access journal mBio. They stated that:
“Pathogen host shifts represent a major source of new infectious diseases. Here we provide evidence that a pollen-borne plant virus, tobacco ringspot virus (TRSV), also replicates in honeybees and that the virus systemically invades and replicates in different body parts. In addition, the virus was detected inside the body of parasitic Varroa mites, which consume bee hemolymph, suggesting that Varroa mites may play a role in facilitating the spread of the virus in bee colonies. This study represents the first evidence that honeybees exposed to virus-contaminated pollen could also be infected and raises awareness of potential risks of new viral disease emergence due to host shift events. About 5% of known plant viruses are pollen transmitted, and these are potential sources of future host-jumping viruses. The findings from this study showcase the need for increased surveillance for potential host-jumping events as an integrated part of insect pollinator management programs”.
This paper has caused all sorts of excitement, as well as coming to some possibly misleading conclusions, and leading to quite a lot of uninformed speculation – so I think it is as well to explore quite carefully what they did.
Adrian first:
“This paper reports that tobacco ringspot nepovirus, a well-known virus of plants, replicates in honeybees. TRSV, first identified nearly a century ago in the USA, has a wide range of plant hosts, and is spread in pollen and seed, and also by many unrelated vectors, not only root-feeding nematodes, like other nepoviruses, but also insects and mites.
This report tells us that TRSV virions have been isolated from bees, and that their gene sequences are closely similar to those of TRSV. Convincingly, biochemical tests showed that there were replication intermediates of the TRSV genome in the bees, so the virus had not merely contaminated the bees when they fed on the honey and pollen of infected plants, but had seemingly multiplied in them.
However, there are several gaps in this story. Surprisingly, it seems that no tests were done to show whether the virus isolated from bees infected known plant hosts of TRSV; perhaps this crucial evidence will be in the sequel. Furthermore, the reported sequences of the virus represented only around 20% of the RNA1 of typical nepoviruses, and around 50% of their RNA2, so there is still the possibility of further genetic surprises when the genome sequence is completed.
Although TRSV is a well-known and long studied virus with many distinct symptom variants, there are relatively few of its gene sequences in Genbank. When these are compared with those of the ‘bee TRSV’ it is obvious that more sequences will be required to sort out exactly where this virus clusters; differences in the homology patterns of the RNAs 1 and 2 suggest that recombination or reassortment is active among nepoviruses.
The gene sequencing revolution is, to paraphrase Pliny the Elder, revealing that in virology the only certainty is that nothing is certain.”
Me next:
I have the same reservations as Adrian: the TRSV sequence isolated from bees and from Varroa mites is only partial (~30%); thus, it is by no means certain that the whole genome is colinear with genomes of established TRSVs or of other nepoviruses, although they assume that it is – with some justification, possibly, as sequence identities of up to 96% were found in Genbank for the putative CP fragment sequence. They could isolate particles: why, then, in this metagenomic and NGS age, could they not sequence the whole thing??
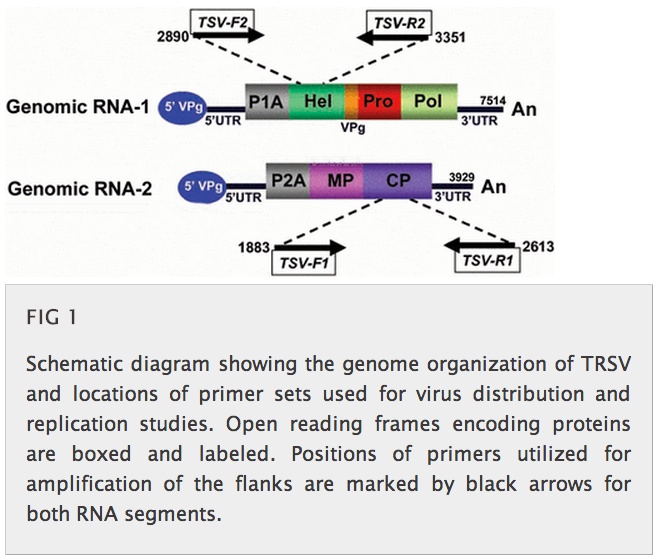
Another reservation I have concerns their methodology. First, while I was impressed that they did strand-specific PCR to show both the presence of viral (+ strand) and of replicative form (- strand) RNA in bee and mite tissue (see their Figure above), and did in situ hybridisation to show (-) strand presence in mites, they did not do something very simple that could have shown the same thing, AND given them genome-length +/- strand RNA to play with.
I refer, of course, to dsRNA isolation, which is a very easy and extremely clean technique that can be used to get full-length dsRNAs for many (+) strand RNA viruses from plant or insect tissues. Moreover, the simple fact of isolating dsRNA forms for a single-strand RNA virus is indicative that replication is occurring – and was used by our group as long ago as 1988 (C Williamson, PhD Thesis, UCT) to isolate full-length ~10 kb dsRNA for two aphid picorna-like viruses.
This means they could have had clear and simple evidence via dsRNA extraction of ALL of the coinfecting viruses present – without all of the expense of total cDNA sequencing. And sampled more hives….
Second, I have to echo Adrian: “…no tests were done to show whether the virus isolated from bees infected known plant hosts of TRSV”. Why ever not? I am afraid that if I were a referee, I would have insisted on this: they did enough other work, after all, that this would not exactly have been an onerous requirement!
And here’s another thing: the authors say, in their Discussion,
“The finding from this study illustrates the complexity of relationships between plant pathogens and the pollinating insects and emphasizes the need for surveillance for potential host-jumping events as an integrated part of insect pollinator conservation.”
Ummmm…no, it doesn’t. This is overstating the significance of their results by an order of magnitude at least. They have simply illustrated that ONE species of honeybee may be infectable by ONE species of plant virus, and that this is ASSOCIATED with “weak” colonies. Moreover, while the presence of TRSV was apparently associated with four weak colonies (out of only ten surveyed), it is quite possible that this is simply the emergence of a commensal-type infection against a background of known bee viruses, and in particular Israeli acute paralysis virus which was found in the same colonies (and blogged on here). The authors also seem to take it as a given that the “emergence” of TRSV into bees is a recent jump – when it may not be recent at all. Their statement in the abstract that
“The tree topology indicated that the TRSVs from arthropod hosts shared a common ancestor with those from plant hosts and subsequently evolved as a distinct lineage after transkingdom host alteration”
is pretty much unsubstantiated, in the absence of any investigation of the lineage in plants or in other bee colonies. Further, they say that
“This study represents a unique example of viruses with host ranges spanning both the plant and animal kingdoms. “
Ummmmm….it doesn’t really do that, either: there are a LOT of arboviruses, with quite a few of them infecting insects and plants. Here, for example, is an illustration from my teaching material of why it is that I think that viruses of insects and plants are an underappreciated evolutionary link for later evolution of viruses that got into mammals.

Transkingdom viruses
I note that bunyaviruses, rhabdoviruses, reoviruses and (not shown) picorna-like viruses appear linked by the fact that insects have possibly the most diverse representatives of these families, which may indicate that these originated in insects. Which were the first complex animals to crawl out of the oceans, to join…plants on dry land? Which explains how plants link up with the far more closely related (in evolutionary terms) insects and vertebrates: plants and insects were alone together for a long, long time before things with spines lurched up out of the water to join them. So were their viruses.
I also said the following in the material there:
“A complicating factor in the picture of viruses co-evolving with their hosts over millennia is the fact that viruses apparently can – and obviously do – make big jumps in hosts every now and then. It seems obvious, for example, that arthropods are almost certainly the original source for a number of virus families infecting insects and mammals – such as the Flaviviridae – and probably also of viruses infecting insects and other animals and plants – such as the Rhabdoviridae and Reoviridae – as well (see also here). For example, picornaviruses of mammals are very similar structurally and genetically to a large number of small RNA viruses of insects and to at least two plant viruses, and – as the insect viruses are more diverse than the mammalian viruses – probably had their origin in some insect that adapted to feed on mammals (or plants) at some distant point in evolutionary time.”
Now quite a lot of interest has been shown in this paper in the blogosphere, and there have been quite a few conclusions drawn from the results that I think are largely unsubstantiated. For example, this Sci Am blog claims
“When HIV jumped from chimpanzees to humans sometime in the early 1900s, it crossed a gulf spanning several million years of evolution. But tobacco ringspot virus, scientists announced last week, has made a jump that defies credulity. It has crossed a yawning chasm ~1.6 billion years wide.”
Again, ummmm…in light of the discussion above, not necessarily! I am of the opinion that picorna-like viruses were shared between insects and plants, and then between insects and animals, hundreds of millions of years ago. And TRSV is a nepovirus – and nepoviruses look like nothing more or less than a picornavirus with a divided genome.
I think TRSV represents something coming back into insects. And I think we will probably find a lot more of them.






