Human warts in all their forms – cutaneous, verrucous and genital growths and lesions – have been known since antiquity, and it was known since at least 1823 that at least some were infectious. Experiments done with human volunteers in the 1890s confirmed this, when it was shown that transplanting wart tissue resulted in typical disease. As early as 1908, it was shown by a G Ciuffo that “verrucae volgare” – common warts – could be transmitted via a cell-free filtrate. However, it was Richard E Shope who first showed that a papillomavirus was associated with animal tumours. A useful review from 1931 on “Infectious oral papillomatosis of dogs” by DeMonbreun and the Ernest Goodpasture of egg culture fame covers the early history of the investigation of human disease as well as of animal papillomas very well, so we will not cover this further.
In light of later findings of the involvement of papillomaviruses, it was a prescient although premature observation by an Italian physician named Rigatoni-Stern in 1842 that cervical cancer appeared to be sexually transmitted, given that it occurred in married women, widows and prostitutes, but rarely in virgins and nuns.
Although papillomaviruses had been implicated as the first viruses known to cause a cancer in mammals as early as the 1930s, and the structurally very similar papovaviruses were similarly implicated in the late 1950s, it was only in 1972 that Stefania Jabłońska proposed that a human papillomavirus (HPV; then called a papovavirus) was involved with the rare hereditary skin cancer called epidermodysplasia verruciformis.
Meanwhile Harald zur Hausen had been investigating since 1974 the involvement of HPV in genital warts (condyloma accuminata) and squamous cell carcinomas, using DNA-based techniques such as hybridisation. The rarely malignant condylomas had been shown to contain papillomavirus particles in some cases in 1968, with a better association in 1970; however, cross-hybridisation studies by zur Hausen’s group on DNA of these and common wart viruses showed no relationship despite their very similar morphologies.

Virus particles from genital warts (6 &7) and a common skin wart (8). Reproduced from Brit. J. vener. Dis., JD Oriel and JD Almeida, 46, 37-42, 1970 with permission from BMJ Publishing Group Ltd.
Zur Hausen speculated on the role of HPVs in squamous cell carcinomas in 1977; Gérard Orth and Jabłońska and colleagues went on to define the “…Risk of Malignant Conversion Associated with the Type of Human Papillomavirus Involved in Epidermodysplasia Verruciformis” in 1979.
Because this was the new era of cloning and sequencing of DNA, the zur Hausen group and others went on to isolate and characterise a number of new HPVs associated with genital cancers and other lesions in the early 1980s. In particular, they showed that HPV types 16 and 18 could be found both as free virus in cervical cell sample biopsies and integrated into the cell genomes of cell lines derived from cervical cancers. A major finding in 1987 was that the legendary HeLa cell line – derived from a malignant cervical tumour from a Henrietta Lacks in 1951 – contains multiple copies of the HPV-18 genome. The first HPV genome sequence (of type 1b) was obtained in 1982; the first genital type (6b, from condylomas) in 1983, and the first high-risk cancer virus (type 16) in 1985.
Later work involving large international surveys showed by 1995 that 99.7% of cervical cancers contained DNA from so-called “high risk” HPVs, leading to the conclusion that these were the necessary cause of cervical cancer, and that around 70% of these cancers were caused by HPVs 16 and 18. Since then, HPVs have been found in more than 80% of anal cancers, 70% of vulval and 40% of vaginal cancers, around half of all penile cancers, and in roughly 20% of head and neck cancers. If 16% of cancers are due to infection, and HPVs cause or are implicated in 30% of these, then they are a significant cause of cancers worldwide.
Harald zur Hausen was awarded a half share of the 2008 Nobel Prize in Physiology or Medicine “for his discovery of human papilloma viruses [sic] causing cervical cancer”. I blogged on this at the time, here.
Work on vaccines against papillomaviruses (PVs) started early, after demonstrations presumably in the 1930s that domestic rabbits inoculated with the cottontail rabbit PV (CRPV) could become immune to reinoculation after recovery, and in 1962 that a “…formalin-treated suspension of bovine papilloma tissue” provided protection against challenge, but was not therapeutic. However, progress was stymied by the fact that it proved impossible to culture any of the PVs, and challenge material had to be made from infected animal tissue, even though it had been shown that isolated viral DNA was infectious.
This changed after the advent of molecular cloning, when whole viral genomes could be prepared in bacteria. Model systems for use in PV vaccine research by 1986 included cattle and bovine PVs, rabbits and CRPV and rabbit oral PV, and dogs and canine oral PV. It had also been demonstrated that the L1 major structural protein of type 1 BPV produced in recombinant bacteria was protective against viral challenge in calves. Jarrett and colleagues demonstrated, in 1991 and 1993 respectively, that they had achieved prophylactic and therapeutic immunisation against cutaneous (ie: skin; caused by BPV-2) and then mucosal (respiratory tract; BPV-4) bovine PVs, using E coli-produced proteins. L1 and L2 proteins were protective against BPV-2, while L2 was protective against BPV-4 infection. They suggested BPV-4 was a good model for HPV-16 given its mucosal tropism.
By the early 1990s several groups had demonstrated that it was possible to make PV virus-like particles (VLPs) by expression in eukaryotic systems such as yeast or animal cells of the L1 major virion protein either alone, or together with the minor protein L2. In 1991 Ian Frazer’s group showed that expression of HPV-16 L1 and L2 together but not separately in animal cells via recombinant vaccinia virus, resulted in 40 nm particles resembling the virion being made. In 1992 John Schiller’s lab showed VLP formation by L1 alone, with both BPV-1 and HPV-16 L1 genes expressed in insect cells via a baculovirus vector. In 1993 came the demonstration that expression of the plantar wart-causing HPV-1 L1 gene alone and L1 and L2 genes together in animal cells via vaccinia virus, as well as of the genital wart-causing HPV-11 L1 expressed in insect cells, resulted in VLP formation. By 1995, it had been shown that immunisation of rabbits with CRPV L1-only or L1+L2 VLPs, and of dogs with canine oral PV L1 VLPs, protected completely against viral challenge.
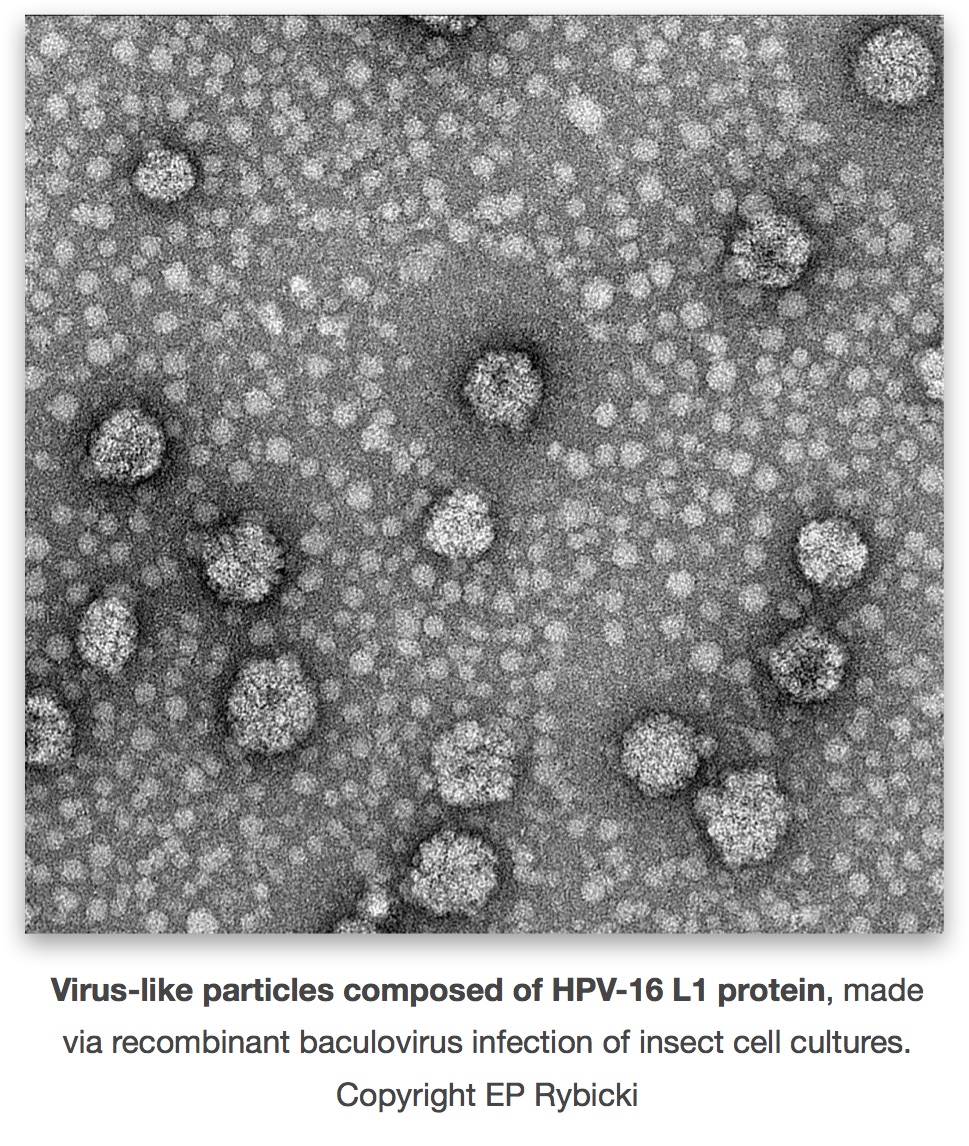
This groundwork made it possible for Merck and GlaxoSmithKline to develop and to push through to human trial and licensure, two independent VLP-based vaccines. Merck’s vaccine – Gardasil – is quadrivalent, consisting of a mixture of VLPs made in recombinant yeasts from expression of L1 genes of HPV types 6 and 11, to protect against genital warts, and types 16 and 18, for cervical lesions and cancer. GSK’s offering – Cervarix – is a bivalent HPV-16 and -18 vaccine only, consisting of VLPs made via recombinant baculoviruses in insect cell culture. These are only the second anti-cancer vaccines on offer, and have gone on to blockbuster status within months of their release: Gardasil was licenced in June 2006, and Cervarix in October 2009.
Both appear to protect very well against infection with the types specified, but not to affect established infections. Their long-term efficacy against cervical cancer is still to be established, although Gardasil has certainly lessened the incidence of genital warts in Australia post introduction in 2007. There is now also a VLP-based vaccine for canine oral PV.
